Monohybrid crossing called one in which the parent forms differ in only one pair of alternative or contrasting characters.
For example, the father plant bears purple flowers and the mother plant bears white flowers, or vice versa.
Before crossing, it is necessary to make sure that the selected characteristics of the parental forms are contrasting in a number of generations, i.e., during self-pollination or close inbreeding, each of the selected characters is persistently inherited. Related organisms that reproduce the same hereditarily constant characteristics in a number of generations are usually called a line.
In plants with hermaphroditic flowers, during artificial hybridization, before pollination, the flowers of the mother plant are castrated, removing the anthers before they are ripe. Same-sex female flowers of cross-pollinators are placed in isolators in advance. When the stigmas ripen, pollen collected from the flowers of the parent plant is applied to them.
When plants, for example, peas, are crossed, the seeds that ripen in the bean on the mother plant in the year of crossing are already first-generation hybrids (F 1). From these sown seeds, hybrid plants of the first generation will grow, and in the beans of these plants, as a result of self-pollination, seeds with embryos of the second generation (F 2) will develop. If the mother plant had flowers, for example, purple, and the father plant had white flowers, then the flowers of the F 1 hybrid plants are all purple; plants with white flowers do not appear among them.
When crossing plants that differ in seed color (yellow and green), the hybrid seeds on the mother plant turn out to be only yellow in the year of crossing. If the original plants differed in the shape of the seeds (smooth - wrinkled), then the F 1 hybrid seeds on the mother plant turn out to be only smooth.
Consequently, in a first-generation hybrid, only one of each pair of alternative characters develops. The second sign seems to disappear and does not appear. G. Mendel called this phenomenon of the predominance of a hybrid trait of one of the parents dominance. A trait that appears in a first-generation hybrid and suppresses the development of another trait was called dominant; opposite, i.e. suppressed, sign - recessive. Mendel defines dominant and recessive characters as follows: “Characters that pass into hybrid compounds completely unchanged or almost unchanged and thereby represent the characteristics of hybrids will be designated as dominant, and those that become latent during hybridization will be designated as recessive.”
Law of Dominance- Mendel's first law - is also called the law of uniformity of first-generation hybrids, since all individuals of the first generation have the same manifestation of the trait.
If a hybrid of the first generation, obtained, for example, from crossing two forms of peas that differ in flower color, has the opportunity to self-pollinate, then in the next generation, i.e. in F 2, plants with characteristics of both parents appear. This phenomenon is called splitting. In F2, splitting is observed in a completely definite quantitative ratio, namely: on average, 3/4 of the total number of plants bear purple flowers and only 1/4 have white flowers, i.e., the ratio of the number of plants with a dominant trait to the number of plants with a recessive trait turns out to be equal to 3: 1. Consequently, the recessive trait in the first generation hybrid did not disappear, but was only suppressed and appeared in the second hybrid generation.
Each plant from F 2 with white flowers, when self-pollinated in the next generations - F 3 and F 4, etc., produces plants with only white flowers. Plants with purple flowers behave differently. Only 1/3 of them, when self-pollinated, produce plants with only purple flowers in F 3 and subsequent generations, and the remaining 2/3 again produce plants of both types in the ratio: 3 plants with purple flowers and 1 with white flowers.
Consequently, the class of plants F 3 with a dominant trait breaks down according to its hereditary inclinations in the ratio 1: 2, and all plants of the second generation give a ratio when splitting into one pair of hereditary inclinations 1: 2: 1. The concept of a class here and in what follows is used in the sense of a group descendants similar in the studied trait or hereditary inclinations.
Everything stated in relation to the inheritance of flower color is also applicable to the inheritance of any other pair of alternative characters, however, under certain conditions, which will be discussed below.
Thus, when studying the inheritance of the smooth or wrinkled form of seeds from 253 hybrid self-pollinating plants F 1, Mendel obtained 7324 seeds in F 2, of which 5474 were smooth, 1850 were wrinkled. If the ratio of 3: 1 is correct, then with a total number of seeds of 7324 theoretically, the following distribution should be expected: 1/4 of the seeds (i.e. 7324 X 1/4 = 1831) should have a recessive trait (wrinkled), and 3/4 (i.e. 7324 X 3/4 = 5493) should have a dominant one (smooth). In Mendel's experiments, figures were obtained that were very close to theoretical ones.
In another experiment, where the color of seeds (yellow or green) was taken into account, Mendel obtained the following ratio in F2: out of 8023 seeds, 6022 were yellow and 2001 were green, i.e., again, a ratio very close to 3: 1.
However, Mendel repeatedly emphasized that these ratios reflect only average values; with a small number of individuals, the number of plants with alternative traits in F2 will fluctuate due to random reasons.
The data suggest that there is obvious variation among individual plants in the ratio of seed classes, but the total results in a ratio close to the expected 3:1 distribution. Experimental data gave a ratio very close to this - 355: 123.
So, having carried out monohybrid crosses, Mendel established the following patterns of inheritance.
1. In first-generation hybrids, only one of a pair of alternative characters manifests itself - the dominant one, while the recessive one does not appear. This phenomenon was called dominance, and later - Mendel's first law, or the law of uniformity of first-generation hybrids.
2. In the second generation of hybrids, individuals with both a dominant trait and a recessive one appear, the ratio of the former to the latter is on average 3:1. In 1900, G. de Vries proposed calling this phenomenon the law of splitting, and subsequently it was called Mendel’s second law. Descendants with a recessive trait remain constant in subsequent generations during self-pollination.
3. Among 3/4 of the plants of the second generation with a dominant trait, 2/4 of the total number of descendants turn out to be hybrid; when self-pollinated, they again give splitting in F 3 in a ratio of 3: 1, and only 2/4 remains constant in subsequent generations, like the original parental forms and plants from F 2 with a recessive trait. Consequently, in F 2, half of the plants are hybrid, and half are “pure”, constantly retaining their parental characteristics. Thus, the descendants of F 1 hybrids according to these hereditary characteristics are split in the ratio 1: 2: 1. Mendel formulated this as follows: “Hybrids according to two different characteristics form seeds, of which half give rise to a newly hybrid form, while the other produces plants that remain constant and contain dominant and recessive traits in equal proportions.”
As we have seen, in F 2 one should distinguish, firstly, splitting according to the external manifestation of characteristics, which is expressed by the ratio 3: 1, and, secondly, according to hereditary potentials, inclinations, expressed by the ratio 1: 2: 1. The first type of splitting is called splitting according to phenotype, i.e., according to the external manifestation of characteristics, the second type - according to the genotype, i.e., according to hereditary inclinations. The terms “phenotype” and “genotype” were introduced in 1903 by V. Johannsen.
By genotype we mean the totality of hereditary inclinations that an organism possesses. A phenotype is a set of properties and characteristics of an organism that are the result of the interaction of an individual’s genotype and the environment.
Mendel was the first to use the symbolic designation of the genotype, where the hereditary factors that determine paired alternative traits were designated by letters of the Latin alphabet. In 1902, V. Bateson proposed to call such a pair of alternative characters an allelomorphic pair, and the pairing of characters - allelomorphism. In 1926, V. Johannsen proposed the term “ allelomorphism"replace with a shorter one - "allelism", and call the individual factor of one pair "allele". The terms “dominant” or “recessive allele” began to mean an alternative state of the same gene. The same meaning was given to the previous designations firmly established in genetics - “dominant gene” and “recessive gene”.
Mendel designated the dominant allele, which determines the trait of yellow color of seeds, with a capital letter A, and the recessive allele, which determines the trait of green color, with a lowercase letter a; the genotype of the dominant form is AA, and the recessive form is aa; hybrid F 1 - Aa. In this case, the descendants of the hybrids show splitting in F 2, corresponding to the formula: 1AA: 2Aa: 1aa.
This symbolism of factors of paired characteristics, used to display the patterns of segregation in the offspring of hybrids, is a kind of “algebra” of genetics, since instead of letter expressions, various genes and alleles corresponding to them can be substituted.
In 1902, V. Bateson proposed to call the constant forms AA and aa, which do not produce cleavage in subsequent generations, homozygous, and the forms Aa that give cleavage, heterozygous. These terms are widely used in genetics. They come from the term “zygote”, i.e. fertilized egg.
During the process of fertilization, the combination of male and female gametes A and A or a and a that are identical in factors gives a homozygote, or homozygous individual AA or aa, and the combination of gametes that differ in factors A and a produces a heterozygote, or heterozygous individual Aa.
As we have seen, in the first generation hybrids the recessive allele a, although not manifested, is not mixed with the dominant A, and in the second generation both alleles again appear in a “pure” form. This phenomenon can be explained only on the basis of the assumption that the first-generation hybrid Aa forms not hybrid, but pure gametes, and the indicated alleles appear in different gametes. Gametes carrying alleles A and a are formed in equal numbers; Based on this, the splitting according to the genotype 1:2:1 becomes clear.
The non-mixing of the alleles of each pair of alternative characters in the gametes of a hybrid organism is called the phenomenon of gamete purity; the second is based on the cytological mechanism of meiosis.
Mendel discovered an interesting phenomenon of changes in the ratio of homozygotes and heterozygotes in a series of successive generations of a hybrid during self-pollination. If we assume that, on average, all plants in a series of generations have the same fertility and viability and take into account the fact that hybrids split in the second generation in a certain numerical ratio (namely 2 homozygous and 2 heterozygous plants for a given trait), then in subsequent generations during self-pollination In all plants, the number of homozygotes will increase, and the number of heterozygotes will decrease.
For simplicity, Mendel assumed that each plant, when self-pollinating, produces four seeds in each generation. In this case, the ratio of homozygotes and heterozygotes for one pair of traits will change over a number of generations. In the tenth generation, according to Mendel’s calculations, for every 2048 plants that should arise at the given reproduction rate, 1023 will be constant - homozygous for a dominant trait, 1023 - homozygous for a recessive trait, and only 2 hybrid, i.e. heterozygous forms will appear. These calculations of Mendel, as we will see later, are important for understanding the genetic basis of selection and gene dynamics in a population.
So far we have talked about one pair of alternative traits, one of which is dominant, the other recessive. Mendel studied seven pairs of the following characters in peas:
1) the seeds are smooth and wrinkled, or angular, 2) the cotyledons are yellow and green, 3) the color of the seed coat is gray-brown and white, 4) the shape of the bean is convex and constricted, 5) the color of the mature bean is yellow and green, 6) the arrangement of flowers axillary and apical, 7) plant stem is high and low. For each of these seven pairs of traits separately in F2, phenotypic segregation was observed in an average ratio of 3:1. Expressing this as a percentage, we can say that in F 2 there were about 75% of plants with dominant and about 25% with recessive traits, or dominant homozygotes - 25%, heterozygotes - 50% and recessive homozygotes - 25%.
The true nature of the pairing of characters remained unknown to Mendel. He assumed that germ cells carry one hereditary inclination, which are united in pairs during fertilization. Now these inclinations, or factors carried by germ cells, are called genes. For now, by the term “gene” we will understand a unit of heredity that determines the development of a separate trait, or property, of an organism. As we deepen the analysis of the phenomena of heredity and the mechanism of inheritance, our understanding of the nature of the gene will expand. Genes, as we have indicated, determine the development of the characteristics of the organism and must be associated with the material structures of the germ cells. Obviously, in the time of Mendel, only guesses could be made on this score, since the structure and development of germ cells had not yet been studied.
What determines the pairing of genes, the purity of gametes and the strict distribution of genes in the offspring, which determines splitting in a certain numerical ratio? After we became acquainted in previous chapters with the development of germ cells and the formation of gametes, it is easy for us to connect the phenomenon of character splitting observed by Mendel with the behavior of chromosomes: their pairing, the divergence of homologous chromosomes in meiosis and their reunification during the process of fertilization.
Let us assume that in the somatic cells of a plant there is only one pair of homologous chromosomes, and the gene that determines the trait of purple flower color, designated A, is located in each of these chromosomes in the parent plant. Then the somatic cells of a homozygous plant, possessing a dominant flower color gene, must carry two dominant AA alleles, since in somatic cells each of the homologous chromosomes is represented in double quantity. Accordingly, the cells of another parent plant with white flowers have a homozygous recessive allele for white color, i.e. aa.
As a result of meiosis, the number of chromosomes in each gamete is halved and only one chromosome from a pair remains. Consequently, the gene located on this chromosome is presented in the gamete in the form of alleles A or a. As a result of fertilization, the chromosome pairing is restored in the hybrid zygote, and the formula of the hybrid will be exactly the same as Mendel wrote it - Aa. During the development of germ cells in a hybrid organism in meiosis, the chromosomes of a given pair will separate into different daughter cells. Then male and female gametes will carry one of the alleles of the gene: A or a. Such gametes, both male and female, will be formed in equal numbers. During fertilization, the types can combine with equal probability to form four types of zygotes.
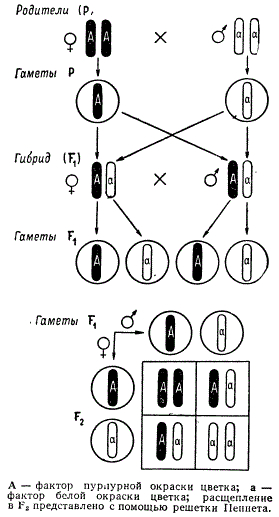
To facilitate the calculation of combinations of different types of gametes, the English geneticist R. Punnett proposed recording in the form of a lattice, which entered the literature as the Punnett lattice. To the left of the grid, female gametes are indicated vertically, and male gametes are indicated horizontally. The resulting combinations of gametes fit into the squares of the lattice. These combinations correspond to the genotypes of the zygotes. The Punnett grid is especially useful for analyzing the inheritance of traits in complex hybrids.
Consideration of the above diagram shows that cleavage according to the 3:1 phenotype and 1:2:1 genotype can be carried out only under certain conditions. Firstly, there must be an equally probable formation in meiosis of both types of gametes, i.e. gametes carrying the A allele and gametes with the a allele. Secondly, there must be an equally probable meeting and combination of these gametes during fertilization. Both of these conditions, as we know from the previous chapter, are provided with greater accuracy with a larger number of observations. In the first division of meiosis, the number of chromosomes is reduced with the divergence of homologous partners to the poles, and the probability of chromosomes moving to one or another pole is the same, as is the probability of maturation and development of gametes of all varieties. A large number of gametes also ensures that they are equally likely to meet during fertilization. To these conditions it is necessary to add one more thing - equal survival of all types of zygotes and the individuals developing from them.
When analyzing monohybrid crosses, we did not pay attention to which of the plants was maternal and which was paternal. Does the fact that the mother plant will carry a dominant trait, and the male plant will carry a recessive trait, and vice versa, affect the properties of the hybrid and the nature of segregation in its offspring? Even before Mendel, hybridizers noticed that the direction of crossing usually does not affect the characteristics of the hybrid. This gave reason to assume equal participation of the female and male sexes in the transmission of hereditary factors. Mendel confirmed these observations. A dominant trait appears in a hybrid regardless of whether the trait is contributed by the maternal or paternal plant. For the pea traits, the inheritance of which Mendel studied, this position was correct.
But it must be said that sometimes there are differences in the transmission of hereditary properties from the maternal or paternal body; we will get acquainted with this phenomenon later. Therefore, it is still customary to indicate the direction of crossing. Crossings of two forms with each other in two different directions are called reciprocal. So, when crossing two forms P 1 and P 2 in one direction, P 1 acts as the maternal form, P 2 as the paternal form (P 1 XP 2), and in the second - P 2 as the maternal, P 1 as the paternal (P 2 XP 1).
If you find an error, please highlight a piece of text and click Ctrl+Enter.
N. Mendel formulated the patterns of inheritance of traits based on an analysis of the results of monohybrid crossing and called them rules, later they became known as laws.
As it turned out, when crossing two pure lines of peas with yellow (AA) and green (aa) seeds in the first generation (F 1), all hybrid seeds were yellow. Consequently, the trait of yellow seed color was dominant:
R: AA x aa
Similar results were obtained by Mendel when analyzing the inheritance of other six pairs of traits. Based on this, Mendel formulated dominance rule, or first law : in a monohybrid crossing, all offspring in the first generation are characterized by uniformity in phenotype and genotype, named later law of uniformity of first generation hybrids:
When crossing homozygous individuals that differ in one or several pairs of alternative traits, the unity of all first-generation hybrids is observed, both in phenotype and genotype.
From seeds obtained from first-generation hybrids, Mendel grew plants, which he either crossed with each other or allowed to self-pollinate. Among the descendants (F 2), a split was revealed: in the second generation there were both yellow and green seeds.
R: Aa x Aa In total, Mendel received 6022 yellow and 2001 green seeds (numerical G: A a A a ratio approximately 3:1). The same numerical ratios were F2: AA, Aa, Aa, aa and were obtained for the other six pairs of pea traits studied by Mendel. Eventually Mendel's second law is formulated as follows:
When monohybrid crossing heterozygous individuals (first generation hybrids) in the second generation, a splitting ratio of 3:1 in phenotype and 1:2:1 in genotype is observed.
Rice. 1.
75% of all seeds (AA, 2Aa) in the second generation, having one or two dominant alleles, were yellow in color and 25% (aa) were green. The fact that recessive traits appear in the second generation indicates that these traits, as well as their controlled genes, do not disappear, are not mixed with dominant traits in a hybrid organism, their activity is simply suppressed by the action of dominant genes.
A dominant gene does not always completely suppress the action of a recessive gene. In this case, the first generation hybrids do not reproduce the characteristics of their parents - there is intermediate nature of inheritance. In the second generation, dominant homo- and heterozygotes differ in phenotype and genotype equally (1:2:1).
For example, when crossing homozygous night beauty plants with red (AA) and white (aa) flowers, the first generation is obtained with pink flowers (intermediate inheritance). In the second generation, splitting by phenotype corresponds to splitting by genotype: one part of the plants has red flowers, two parts have pink flowers, and one part has white flowers:
R: AA x aa R: Aa x Aa
G: A a G: A a A a
F 1: Aa F 2: AA, Aa, Aa, aa.
To explain the essence of the phenomena of uniformity of hybrids of the first generation and splitting of characters in hybrids of the second generation in 1902, W. Bateson put forward hypothesis of "purity" of gametes, which can be briefly summarized as follows:
- 1) in a hybrid organism, genes do not hybridize (do not mix), but are in a pure allelic state;
- 2) during the process of meiosis, only one gene from an allelic pair enters the gamete.
The gamete purity hypothesis establishes that the laws of segregation are a consequence of a random combination of gametes carrying different genes. However, the general result turns out to be natural, since a statistical pattern is manifested here, determined by a large number of equally probable meetings of gametes. Thus, splitting during monohybrid crossing of heterozygous organisms 3:1 in the case of complete dominance or 1:2:1 in the case of incomplete dominance should be considered as a biological pattern based on statistical data.

Rice. 2. Cytological basis of monohybrid crossing and the “gamete purity” hypothesis
The cytological basis of the hypothesis of gamete purity and the first two laws of Mendel are the patterns of divergence of homologous chromosomes and the formation of haploid germ cells in the process of meiosis.
Having worked through these topics, you should be able to:
- Give definitions: gene, dominant trait; recessive trait; allele; homologous chromosomes; monohybrid crossing, crossing over, homozygous and heterozygous organism, independent distribution, complete and incomplete dominance, genotype, phenotype.
- Using the Punnett grid, illustrate crossbreeding for one or two traits and indicate what numerical ratios of genotypes and phenotypes should be expected in the offspring from these crosses.
- Explain the rules of inheritance, segregation, and independent distribution of characters, the discovery of which was Mendel's contribution to genetics.
- Explain how mutations can affect the protein encoded by a particular gene.
- Indicate the possible genotypes of people with blood groups A; IN; AB; ABOUT.
- Give examples of polygenic traits.
- Indicate the chromosomal mechanism of sex determination and types of inheritance of sex-linked genes in mammals, and use this information when solving problems.
- Explain the difference between sex-linked traits and sex-dependent traits; give examples.
- Explain how human genetic diseases such as hemophilia, color blindness, and sickle cell anemia are inherited.
- Name the features of methods of selection of plants and animals.
- Indicate the main directions of biotechnology.
- Be able to solve simple genetic problems using this algorithm:
Algorithm for solving problems
- Determine the dominant and recessive traits based on the results of crossing the first generation (F1) and the second (F2) (according to the conditions of the problem). Enter the letter designations: A - dominant and - recessive.
- Write down the genotype of an individual with a recessive trait or an individual with a known genotype and gametes based on the conditions of the problem.
- Record the genotype of the F1 hybrids.
- Draw up a scheme for the second crossing. Record the gametes of F1 hybrids in a Punnett grid horizontally and vertically.
- Record the genotypes of the offspring in the gamete intersection cells. Determine the ratios of phenotypes in F1.
Task design scheme.
Letter designations:
a) dominant trait _______________
b) recessive trait _______________
Gametes
F1(first generation genotype)
| gametes | ||
| ? | ? |
Punnett grid
| F2 |
|
Phenotype ratio in F2: _____________________________
Answer:_________________________
Examples of solving monohybrid crossing problems.
Task.“There are two children in the Ivanov family: a brown-eyed daughter and a blue-eyed son. The mother of these children is blue-eyed, but her parents had brown eyes. How is eye color inherited in humans? What are the genotypes of all family members? Eye color is a monogenic autosomal trait.”
The eye color trait is controlled by one gene (by condition). The mother of these children is blue-eyed, and her parents had brown eyes. This is only possible if both parents were heterozygous, therefore, brown eyes dominate over blue ones. Thus, grandparents, father and daughter had the genotype (Aa), and mother and son had the genotype aa.
Task."A rooster with a rose-shaped comb was crossed with two hens, also having a rose-shaped comb. The first gave 14 chickens, all with a rose-shaped comb, and the second gave 9 chickens, of which 7 with a rose-shaped and 2 with a leaf-shaped comb. The shape of the comb is a monogenic autosomal trait. What are genotypes of all three parents?
Before determining the genotypes of the parents, it is necessary to find out the nature of inheritance of the comb shape in chickens. When a rooster was crossed with a second hen, 2 chicks with leaf combs were produced. This is possible if the parents are heterozygous; therefore, it can be assumed that the rose-shaped comb in chickens is dominant over the leaf-shaped one. Thus, the genotypes of the rooster and the second hen are Aa.
When crossing the same rooster with the first hen, no splitting was observed, therefore, the first hen was homozygous - AA.
Task.“In a family of brown-eyed, right-handed parents, fraternal twins were born, one of whom is brown-eyed, left-handed, and the other blue-eyed, right-handed. What is the probability of the next child being born similar to his parents?”
The birth of a blue-eyed child to brown-eyed parents indicates the recessiveness of blue eye color, respectively, the birth of a left-handed child to right-handed parents indicates the recessivity of better control of the left hand compared to the right. Let's introduce allele designations: A - brown eyes, a - blue eyes, B - right-handed, c - left-handed. Let's determine the genotypes of parents and children:
| R | AaBv x AaBv |
| F, | A_bb, aaB_ |
A_вв is a phenotypic radical, which shows that this child is left-handed with brown eyes. The genotype of this child may be Aavv, AAvv.
Further solution of this problem is carried out in the traditional way, by constructing a Punnett lattice.
| AB | Av | aB | Av | |
| AB | AABB | AAVv | AaBB | AaVv |
| Av | AAVv | AAbb | AaVv | Aaww |
| aB | AaBB | AaVv | aaBB | AaVv |
| aw | AaVv | Aaww | aaVv | Aaww |
9 variants of descendants that interest us are underlined. There are 16 possible options, so the probability of a child being born similar to their parents is 9/16.
Ivanova T.V., Kalinova G.S., Myagkova A.N. "General Biology". Moscow, "Enlightenment", 2000
- Topic 10. "Monohybrid and dihybrid crossing." §23-24 pp. 63-67
- Topic 11. "Genetics of sex." §28-29 pp. 71-85
- Topic 12. "Mutational and modification variability." §30-31 pp. 85-90
- Topic 13. "Selection." §32-34 pp. 90-97
Monohybrid cross
Phenotype and genotype. Monohybrid called crossing, in which the parent forms differ from each other in one pair of contrasting, alternative characters.
Sign- any feature of an organism, i.e. any individual quality or property by which two individuals can be distinguished. In plants, this is the shape of the corolla (for example, symmetrical-asymmetrical) or its coloring(purple-white), speed of plant maturation (early ripening-late ripening), resistance or susceptibility to disease, etc.
The totality of all signs of an organism, from external to the features of the structure and functioning of cells, tissues and organs, is called phenotype. This term can also be used in relation to one of the alternative characteristics.
The signs and properties of the body appear under the control of hereditary factors, i.e. genes. The set of all genes in an organism is called genotype.
Examples of monohybrid crosses carried out by G. Mendel include crossings of peas with such clearly visible alternative characters as purple and white flowers, yellow and green coloring unripe fruits (beans), smooth and wrinkled surface of seeds, yellow and green coloring, etc.
Uniformity of first generation hybrids (Mendel's first law). When crossing peas with purple and white flowers, Mendel found that all hybrid plants of the first generation (F 1 ) the flowers turned out to be purple. In this case, the white color of the flower did not appear (Fig. 3.1).
Mendel also established that all hybrids F 1 turned out to be uniform (homogeneous) for each of the seven characteristics he studied.
Rice. 3.1. Monohybrid crossing scheme: I - homozygous individuals with a dominant trait; 2 - heterozygous individuals with a dominant or intermediate trait; 3 - homozygous individuals with a recessive trait.
Consequently, in first-generation hybrids, out of a pair of parental alternative traits, only one appears, and the trait of the other parent seems to disappear. The phenomenon of dominance in hybrids F 1 Mendel named the characteristics of one of the parents dominance, and the corresponding sign is dominant. Traits that do not appear in hybrids F 1 he named recessive.
Since all first generation hybrids are uniform, this phenomenon was called by K. Correns Mendel's first laws, or law of uniformity of first generation hybrids, and rule of dominance.
Law of segregation (Mendel's second law) From hybrid pea seeds, Mendel grew plants that self-pollinated, and sowed the resulting seeds again. As a result, the second generation of hybrids, or F 2 hybrids, was obtained. Among the latter, a split in each pair of alternative characters was found in a ratio of approximately 3:1, i.e. three quarters of the plants had dominant characters (purple flowers, yellow seeds, smooth seeds, etc.) and one quarter had recessive characters (white flowers, green seeds, wrinkled seeds, etc.). Consequently, the recessive trait of the F 1 hybrid did not disappear, but was only suppressed and reappeared in the second generation. This generalization was later called Mendel's second law, or the law of splitting.
Homozygous and heterozygous individuals. To find out how the inheritance of traits would occur in the third, fourth and subsequent generations, Mendel raised hybrids of these generations through self-pollination and analyzed the resulting offspring. He found that plants with recessive traits (for example, white flowers) in subsequent generations (F 3 F 4 etc.), reproduce offspring only with white flowers (see Fig. 3.1).
The second generation hybrids, which had dominant traits (for example, purple flowers), behaved differently. Among them, when analyzing the offspring, Mendel discovered two groups of plants, outwardly completely indistinguishable for each specific characteristic.
The first group, constituting 1/3 of the total number of plants with a dominant trait, did not split further, i.e., in all subsequent generations they showed only purple flower color. The remaining 2/3 of the plants of the second generation in F 3 again gave the same splitting as in F 2 that is, for every three plants with purple flowers, one with white flowers appeared.
Individuals that do not produce cleavage in their offspring and retain their characteristics in a “pure” form are called homozygous, and those in whom splitting occurs in their offspring - heterozygous.
Thus, Mendel was the first to establish that plants that are similar in external characteristics can have different hereditary properties.
Allelism. To establish the cause of splitting, and in strictly defined numerical ratios of dominant and recessive characters, it should be remembered that the connection between generations during sexual reproduction is carried out through germ cells (gametes). Obviously, gametes carry material hereditary inclinations, or factors that determine the development of a particular trait. These factors were later called genes.
In the somatic cells of a diploid organism, these inclinations are paired: one is received from the paternal organism, and the other from the maternal. Mendel proposed to designate the dominant hereditary inclinations of the capital letter(For example, A), and the corresponding recessive inclinations of the capital letter (A). A pair of genes that determine alternative traits is called allelomorphic pair, and the phenomenon of pairing itself - allelism.
Each gene has two states - A and a, therefore they form one pair, and each member of the pair is called allele. Thus, genes located in the same loci (sections) of homologous chromosomes and determining the alternative development of the same trait, are called allelic. For example, the purple and white color of a pea flower are dominant and recessive traits, respectively, for two alleles (A And A) one gene. Due to the presence of two alleles, two states of the body are possible: homo- and heterozygous. If an organism contains the same alleles for a particular gene (AA or aa), then it is called homozygous for a given gene (or trait), and if different (Ah)- That heterozygous. Therefore, an allele is a form of existence of a gene.
An example of a three-allelic gene is the gene that determines the human blood group system AB0. There are even more alleles: for the gene that controls the synthesis of human hemoglobin, many dozens of them are known.
Statistical analysis of splitting. Let us present the results of Mendel's experiments on monohybrid crossing of peas in the form of a diagram (Fig. 3.2). Symbols P, F 1 , F 2 etc. denote the parent, 1st and 2nd generation respectively, the multiplication sign indicates crossbreeding, the symbol o* denotes male, and Q - female. From the diagram it is clear that in the parent generation (R) maternal and paternal forms are homozygous for the trait being studied, therefore they produce gametes only with the allele A or only with A.
When fertilized, these gametes form a zygote that has both alleles Ahh- dominant and recessive. As a result, all F 1 hybrids are uniform in a specific trait, since the dominant allele A suppresses the effect of the recessive allele A. During gamete formation, alleles A and a hit them one at a time. Therefore, hybrid organisms are capable of producing two types of gametes carrying alleles A And A, i.e. they are heterozygous.

Rice. 3.2. Inheritance of purple and white color of pea flowers.
To facilitate the calculation of combinations of different types of gametes, the English geneticist R. Punnett proposed recording in the form of a lattice, which entered the literature under the name lattice Penneta(see Fig. 3.2). The female gametes are located vertically on the left, and the male gametes are located on the horizontal top. The resulting combinations of gametes, which correspond to the genotypes of the zygotes, are entered into the grid squares.
When self-pollinating in F 2 the result is a split according to the genotype in relation to 1 AA:2Aa:1aa, i.e. That is, one fourth of the hybrids are homozygous for dominant alleles, half are heterozygous and one fourth are homozygous for recessive alleles. Since genotypes AA And Ahh the same phenotype corresponds - purple color of the flower, the splitting according to the phenotype will be as follows; 3 purple: 1 white. Consequently, cleavage by phenotype does not coincide with cleavage by genotype.
Now it is easy to explain why homozygous white-flowered plants of the second generation with recessive alleles ahh during self-pollination b F 3 They only give to their own kind. Such plants produce one type of gamete, and as a result, no splitting is observed. It is also clear that among purple-flowered 1/3 dominant homozygotes (AA) will also not produce splitting, and 2/3 of heterozygous plants (Ah) will give b F 3 splitting 3:1, as in hybrids F 2
Based on the analysis of the results of monohybrid crosses, not only Mendel’s first and second laws and the rule of dominance were formulated, but also the rule of gamete purity.
Rule of purity of gametes. In monohybrid crossing in case of complete dominance in heterozygous hybrids (Ah) in the first generation, only the dominant allele appears (A); recessive (A) does not get lost and does not mix with the dominant one. IN F 2 both recessive and dominant alleles can appear in their “pure” form. In this case, the alleles not only do not mix, but also do not undergo changes after being together in a hybrid organism. As a result, the gametes produced by such a heterozygote are “pure” in the sense that the gamete A"pure" and does not contain anything from the allele A, and gamete a is “clean” from A. This phenomenon of non-mixing of alleles of a pair of alternative characters in the gametes of a hybrid is called rule of gamete purity. This rule, formulated by W. Bateson, indicates the discreteness of the gene, the immiscibility of alleles with each other and other genes. The cytological basis of the rule of purity of gametes and the law of segregation is that homologous chromosomes and the genes localized in them that control alternative characteristics are distributed among different gametes.
Analyzing crossing. With complete dominance, it is impossible to judge the genotype of an organism by its phenotype, since a dominant homozygote (AA), and heterozygote (Ah) have a phenotypically dominant trait. In order to distinguish a dominant homozygous from a heterozygous, a method called analyzing crossing, i.e., crossing the organism under study with an organism homozygous for recessive alleles. In this case, the recessive form (aa) produces only one type of gamete per allele A, which allows any of the two alleles of the trait under study to appear in the first generation.
For example, in the fruit fly Drosophila, long wings dominate over rudimentary wings. An individual with long wings may be homozygous (LL) or heterozygous (Ll). To establish its genotype, it is necessary to carry out an analytical cross between this fly and a fly homozygous for recessive alleles. If all descendants from this cross have long wings, then an individual with an unknown genotype is homozygous for dominant alleles (LL), If in the first generation there is a split into dominant and recessive forms in a ratio of 1:1, then we can conclude that the organism under study is heterozygous.
Thus, according to the nature of the splitting, it is possible to analyze the genotype of the hybrid, the types of gametes that it forms, and their ratio. Therefore, analytical crossing is a very important technique of genetic analysis and is widely used in genetics and breeding.
Intermediate inheritance
In some organisms, when crossing, the patterns established by G. Mendel are not followed. It is known that in a heterozygous organism, a dominant gene does not always suppress the manifestation of a recessive gene. In some cases, the first generation F1 hybrid does not completely reproduce any of the variants of the parental traits: the expression of the trait is intermediate in nature with a greater or lesser bias towards dominant and recessive states. But all individuals of this generation show uniformity in this trait.
Phenomenonincomplete dominance Let's look at the example of the night beauty plant. When a plant with red flowers is crossed with a plant that has white flowers, hybrids will be formed that will have pink corollas.
The parent plants were homozygous ( AA and aa) for different alleles of the flower color gene, and the hybrids are heterozygous ( Ahh) for this gene. This means that in this case, homozygotes and heterozygotes differ in phenotype.

Rice. 9. Incomplete dominance

Rice. 10. Inheritance of fruit color in strawberries with incomplete dominance
When crossing two heterozygous plants with pink flowers (Aa) from the first generation, in the second generation, splitting occurs according to the genotype 1AA: 2Aa: 1aa, and according to the phenotype, not in the ratio 1: 3, but in the ratio 1 red: 2 pink: 1 white. This means that in the case of incomplete dominance, three classes of second generation hybrids are formed not only by genotype, but also by phenotype: plants with red (1/4), pink (2/4=1/2) and white (1/4) flowers. So, with incomplete dominance in second-generation hybrids, a 1:2:1 split in both genotype and phenotype is observed, and the genotype of an individual is uniquely determined by its phenotype.
With incomplete dominance, second-generation hybrids with dominant and recessive phenotypes are always homozygotes, and those with an intermediate phenotype are heterozygotes.
The basic laws of the transmission of hereditary characteristics from generation to generation were formulated in 1865 by the outstanding Austro-Czech researcher Gregor Mendel. G. Mendel's article, published in a little-read journal, remained unknown for a long time and became widely known only in 1900.
The main research method used by G. Mendel and which formed the basis of modern genetics is called hybridological. Its essence is the crossing (hybridization) of organisms that differ from each other in one or more characteristics.
Monohybrid cross - crossing of forms that differ from each other in one pair of alternative characteristics.
Mendel began his study of inheritance patterns by crossing pea varieties that differed from each other in only one pair of contrasting characters. Thus, when crossing a pea variety that had round seeds with a variety that had wrinkled seeds, all the resulting hybrid seeds were round, regardless of which variety was used as the female parent. By crossing peas with alternative traits, Mendel discovered that the first generation of hybrids were identical. Moreover, the descendants resembled only one of the parents, although they received the genes for this trait from both. The sign of one of the parents seemed to disappear. He named the characteristics that appear in hybrids, dominant (lat. dominas - dominant), and not manifested in hybrids P, - recessive (Latin, recessus - retreat). Mendel designated dominant traits in capital letters of the Latin alphabet, and recessive ones in small letters.
Based on the experimental results, it was found that in the first generation dominance and uniformity of the descendants of the first generation are manifested (however, complete dominance of traits is not always observed). This pattern is called the law, or rule, uniformity of hybrids first generation, or law of domination (Fig. 1).
In 1902, W. Bateson proposed to call the constant forms AA and aa, which do not split in subsequent generations homozygous , and the forms Aa that give splitting are heterozygous .
The above diagram gives a clear idea of Mendel's basic law. Each trait is indivisible and independent, and of a pair of divergent traits, one is dominant (in the diagram - the yellow color of the seeds), and the other is recessive (the green color of the seeds). When crossing, the dominant gene suppresses the recessive gene (green color of seeds), and all offspring end up with yellow seeds. In this case, the recessive gene remains in a latent state when combined with the dominant one. This can be judged from the results of the analysis of the descendants of the second generation.
Rice. 1. Scheme of monohybrid crossing.
For many loci, not one, but several stable states of genes (located in these loci) are known. Such gene states are called multiple allelism , allelomorphism . Different allelomorphs of one locus cause significant differences in the traits that arise under their influence.